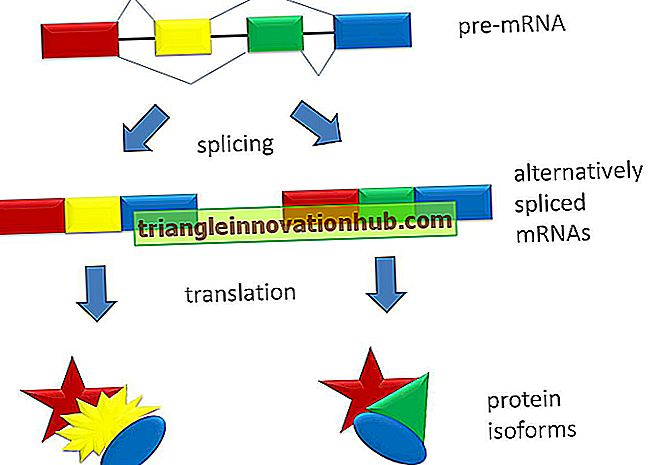
Uwagi na temat fotobiologii w roślinach i jej mechanizmach
Uwagi na temat fotobiologii w roślinach i jej mechanizmach!
Chromoproteina (światłoczuły fotoreceptor, niebieski kompleks białkowy pigmentu znajdujący się w prawie wszystkich roślinach kwitnących (okrytonasienne) został odkryty przez Borthwick i in. (1952) i wyizolowany przez Butlera i wsp. (1959) z etiowanych siewek kukurydzy.
Występuje w dwóch stanach wzajemnie się przestawiających: niebieskiej i żółtej zieleni, P r i P fr . P fr jest związany z błoną komórkową, a P r znajduje się w stanie rozproszonym w cytozolu. Powoduje kiełkowanie nasion, hamuje kwitnienie w SDP i kwitnienie w roślinach długich dni. P r lub P 600 absorbuje światło przy 660 nm i zmienia się na P fr lub P 730 .
Ten ostatni absorbuje światło o długości fali 730 nm i jest szybko zmieniany z powrotem na P r . Ciemność powoduje również tę konwersję, ale jest bardzo powolna. Fitochrom bierze udział w odpowiedzi fotomorfogenetycznej powyżej 550 nm (z wyjątkiem fotonastycznych ruchów fotonastycznych). Jest on niezbędny do kiełkowania nasion niektórych roślin, spoczynku pączków, opadania liści, syntezy giberelin i etylenu, zapobiegania foto-oksydacji, fotoperiodyzmu i morfogenezy.

Forma P r jest stabilna w ciemności, absorbuje czerwone światło o długości 660 nm i szybko zmienia się w P fr . P fr na absorpcję dalekiego czerwonego światła o długości fali 730 nm lub w ciemności zamieniono na P r . To zjawisko zachowawcze jest niezależne od temperatury. Pr stymuluje kwitnienie w roślinach o krótkim dniu. P fr jest niezbędne do kiełkowania nasion.

Mechanizm:
W pewnym okresie fitochrom miał kontrolować reakcje fotomorfogenne poprzez kontrolę metabolizmu związków 2C i 3C (Hendricks, 1964). Później uważano, że działa on poprzez geny (Mohar, 1966) lub zmianę przepuszczalności błony (Hendricks i Borthwick, 1967), działając jako enzym białkowy dla struktury błonowej.
Fondeville i wsp. (1966) zaobserwowali, że fitochrom steruje nycinastic (zależne od światła otwieranie i zamykanie) ruchy ulotek Mimosa. Podobne ruchy sterowane fitochromem zostały później zaobserwowane w juliklissinie Albizzi i aparacie szparkowym. Stwierdzono również, że takie ruchy są związane z napływem i wypływem jonów K + . Dlatego fitochrom powoduje zmiany turbin w komórkach poprzez - zmianę w aktywnym transporcie jonów lub właściwościach membrany.
Fitochrom może wywoływać odpowiedź fotomorfogenetyczną w ciągu kilku sekund (Newman i Briggs, 1972) lub po kilku godzinach. Ten pierwszy nazywa się szybką reakcją fitochromową. Wydaje się, że szybka reakcja fitochromowa jest spowodowana zmianami właściwości błony.
Na przykład wycięte korzenie jęczmienia obecne w zlewce mającej roztwór ATP, Mg2 + i kwasu askorbinowego natychmiast przylegają do powierzchni szkła po wystawieniu na działanie czerwonego światła. Są uwalniane po wystawieniu na dalekie czerwone światło. Wyraźnie widać, że fitochrom znajduje się w błonie komórkowej (Haupt, 1972). Działa poprzez działanie na enzymy związane z błoną.
Operacja fitochromu może również odbywać się za pomocą acetylocholiny lub pokrewnego związku (Jaffe, 1970). Yamamoto i Tezuka (1972) zaproponowali, że fitochrom może działać poprzez regulację NADP + aktywnego w komórkach. Możliwe jest, że zarówno NADP + jak i związki podobne do acetycholiny mogą być związane z niektórymi reakcjami lub półproduktami działania fitochromu.