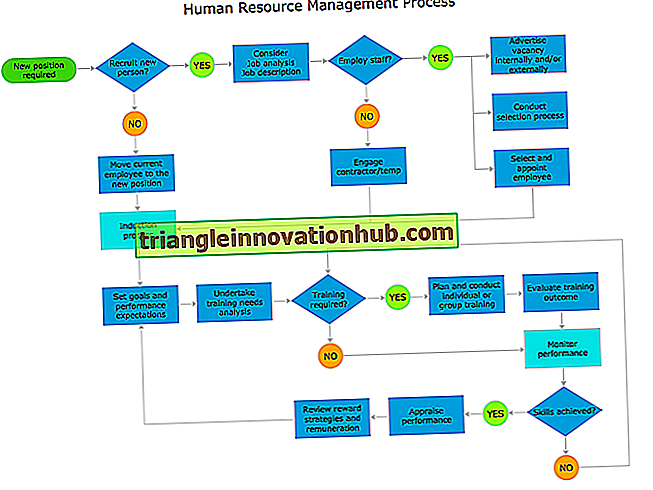
4 Funkcjonalne aspekty ekosystemu
Niektóre z ważnych funkcji ekosystemu są następujące: 1. Przepływ energii w ekosystemie 2. Łańcuch pokarmowy, sieci pokarmowe i piramidy ekologiczne 3. Cykl biogeochemiczny 4. Sukcesja ekologiczna.
Wszystkie ekosystemy utrzymują się w charakterystycznym dynamicznym stanie. Są utrzymywane przez energię, która przepływa przez ich biotyczne składniki i przez obieg materiałów takich jak N, C, H2O wewnątrz i na zewnątrz systemu.
Korzenie ekologiczne w ostatecznej analizie są zorientowane na energię. Ostatecznym źródłem energii jest słońce. Energia słoneczna jest wychwytywana przez autotrofy, przechodzi do heterotrofów relacji producent-konsument, producent-roślinożerca-mięsożerca. Oznacza to, że energia jest przenoszona z jednego poziomu troficznego na drugi z rzędu w postaci łańcucha nazywanego łańcuchem pokarmowym.
Oto funkcjonalne aspekty ekosystemu:
1. Przepływ energii w ekosystemie:
Ekosystemy utrzymują się dzięki energii na rowerze i składnikom odżywczym uzyskanym ze źródeł zewnętrznych. Na pierwszym poziomie troficznym, pierwotni producenci (rośliny, algi i niektóre bakterie) wykorzystują energię słoneczną do produkcji organicznego materiału roślinnego w procesie fotosyntezy.
Roślinożerne zwierzęta, które żywią się wyłącznie roślinami, stanowią drugi poziom troficzny. Drapieżniki, które jedzą roślinożerców, stanowią trzeci poziom troficzny; jeśli obecne są większe drapieżniki, reprezentują one jeszcze wyższe poziomy troficzne.
Organizmy żyjące na kilku poziomach troficznych (na przykład niedźwiedzie grizzly, które jedzą jagody i łososia) są klasyfikowane na najwyższym poziomie troficznym, w którym żyją. Dekompozyty, które obejmują bakterie, grzyby, pleśnie, robaki i owady, rozkładają odpady i martwe organizmy i zwracają składniki odżywcze do gleby.
Średnio około 10% produkcji energii netto na jednym poziomie troficznym przechodzi na kolejny poziom. Procesy, które zmniejszają energię przenoszoną pomiędzy poziomami troficznymi, obejmują oddychanie, wzrost i reprodukcję, defekację i nie-drapieżną śmierć (organizmy, które umierają, ale nie są konsumowane przez konsumentów).
Wartość odżywcza materiału, który jest spożywany, wpływa również na efektywność transferu energii, ponieważ konsumenci mogą przekształcać wysokiej jakości źródła żywności w nową żywą tkankę wydajniej niż źródła żywności o niskiej jakości.
Niski wskaźnik transferu energii między poziomami troficznymi sprawia, że dekompozytorzy są na ogół ważniejsi niż producenci pod względem przepływu energii. Decomposers przetwarzają duże ilości materiału organicznego i zwracają składniki odżywcze do ekosystemu w postaci nieorganicznej, która jest następnie ponownie wykorzystywana przez producentów pierwotnych. Energia nie podlega recyklingowi podczas rozkładu, ale raczej jest uwalniana, głównie w postaci ciepła.
Produktywność ekosystemu:
Wydajność ekosystemu odnosi się do tempa produkcji, tj. Ilości materii organicznej zgromadzonej w jednostkowym przedziale czasowym.
Produktywność ma następujące typy:
(za) Podstawowa wydajność:
Określa się ją jako szybkość, z jaką energia promieniowania jest magazynowana przez aktywność fotosyntezy i chemosyntezy producentów. Ogólna produktywność brutto ekosystemu (GPP) to całkowita ilość materii organicznej wytwarzanej w procesie fotosyntezy.
Wydajność pierwotna netto (NPP) opisuje ilość energii, która pozostaje dostępna dla wzrostu roślin po odjęciu frakcji, którą rośliny wykorzystują do oddychania. Wydajność w ekosystemach lądowych ogólnie wzrasta z temperaturą do około 30 ° C, po czym zmniejsza się i jest dodatnio skorelowana z wilgocią.
Na ziemi wydajność pierwotna jest więc najwyższa w ciepłych, mokrych strefach w tropikach, w których znajdują się biomasy lasów tropikalnych. Natomiast ekosystemy pustynne mają najniższą produktywność, ponieważ ich klimat jest bardzo gorący i suchy.
(b) Drugorzędna wydajność:
Dotyczy konsumentów lub heterotrofów. Jest to wskaźnik zużycia energii na poziomie konsumenckim. Ponieważ konsumenci stosują jedynie pokarm w trakcie oddychania, po prostu zmieniając materiał spożywczy na różne tkanki w całym procesie, wydajność wtórna nie jest klasyfikowana jako ilość brutto i netto. Drugorzędna produktywność faktycznie przemieszcza się z jednego organizmu do innego, tj. Pozostaje mobilna i nie żyje in situ, tak jak pierwotna produktywność.
(c) Wydajność netto:
Odnosi się to do wskaźnika niedoboru materii organicznej niewykorzystywanej przez heterotrofy (konsumentów), tj. Równoważnej pierwotnej produkcji netto minus spożycie przez heterotrofy w jednostce czasu, jako porę roku lub rok itd. Tak więc jest to wskaźnik wzrost biomasy pierwotnych producentów, który został pozostawiony przez konsumentów.

Najprostszym sposobem opisania przepływu energii przez ekosystemy jest łańcuch pokarmowy, w którym energia przechodzi z jednego poziomu troficznego do następnego, bez uwzględniania bardziej złożonych relacji między poszczególnymi gatunkami. Niektóre bardzo proste ekosystemy mogą składać się z łańcucha pokarmowego o zaledwie kilku poziomach troficznych.
Model przepływu energii w kształcie litery Y:
Wiemy, że przepływ energii przez pasożyta można nazwać wypasem łańcucha pokarmowego, a przepływ energii przez detrytusowych konsumentów jako łańcuch pokarmowy detrytus. Partnerzy tych łańcuchów żywnościowych są tak blisko związani, że czasami trudno jest określić ich względny wpływ na rozkład pierwotnej produkcji pierwotnej.

Jak pokazano na rys. 3.2 jedno ramię reprezentuje łańcuch pokarmowy roślinożercy, a drugie łańcuch pokarmowy detrytus. Są ostro rozdzielone. Jednak w warunkach naturalnych nie są one całkowicie odizolowane od siebie.
Na przykład małe martwe zwierzęta, które kiedyś były częścią wypasanego łańcucha pokarmowego, włączają się do łańcucha pokarmowego detrytusu, tak jak twarze wypasanych zwierząt. Ta współzależność reprezentowana w formie figury przypomina literę "Y", dlatego EP Odum (1983) nazwał ją modelem przepływu energii w kształcie litery "Y".
Model w kształcie litery Y jest bardziej realistycznym i praktycznym modelem roboczym niż modele jednokanałowe, ponieważ
za. Potwierdza podstawową strukturę warstwową ekosystemów,
b. Oddziela on łańcuchy pokarmowe wypasu i detrytusu zarówno w czasie, jak iw przestrzeni, oraz
do. Mikro-konsumenci i makro-konsumenci różnią się znacznie relacjami dotyczącymi wielkości metabolizmu.
2. Łańcuch pokarmowy, sieć spożywcza i piramidy ekologiczne:
Łańcuchy pokarmowe:
Łańcuch pokarmowy to szereg populacji, przez które pokarm i zawarta w nim energia przechodzą w ekosystem. Łańcuch pokarmowy jest prosty, jeśli ma tylko jeden poziom troficzny poza rozkładającymi, np. Eichhornia w eutroficznym stawie. Złożony łańcuch pokarmowy ma poziomy troficzne zarówno producenta, jak i konsumenta. Poziomy troficzne są różnymi etapami w przejściu pokarmu.
Istnieją dwa główne typy łańcucha pokarmowego:

(i) Predator lub wypas łańcucha pokarmowego:
Wypasiony łańcuch pokarmowy zaczyna się od fotosyntetycznego wiązania światła, dwutlenku węgla i wody przez rośliny (pierwotni producenci), które produkują cukry i inne cząsteczki organiczne. Po wytworzeniu związki te można stosować do tworzenia różnych typów tkanek roślinnych. Pierwotni konsumenci lub zwierzęta roślinożerne stanowią drugie ogniwo w wypasanym łańcuchu pokarmowym. Zyskują energię poprzez spożywanie głównych producentów.
Wtórni konsumenci lub pierwotni mięsożerne, trzecie ogniwo w łańcuchu, zdobywają energię poprzez spożywanie roślinożerców. Trzecimi konsumentami lub wtórnymi drapieżnikami są zwierzęta, które otrzymują energię organiczną poprzez spożywanie pierwotnych zwierząt mięsożernych.
Przykłady:
1. Trawa → Bydło → Człowiek
2. Trawa → Królik → Lis Wilk → Tygrys
(ii) Łańcuch pokarmowy Detrytus:
Łańcuch pokarmowy detrytus różni się od wypasanego łańcucha pokarmowego na kilka sposobów:
za. Organizmy tworzące są na ogół mniejsze (jak glony, bakterie, grzyby, owady i stonogi)
b. Funkcjonalne role różnych organizmów nie mieszczą się tak zgrabnie w kategoriach, jak troficzne poziomy wypasanego łańcucha pokarmowego.
do. Detrivores żyją w środowiskach (takich jak gleba) bogatych w rozproszone cząstki żywności. W rezultacie rozkładniki są mniej ruchliwe niż zwierzęta roślinożerne lub mięsożerne.
re. Dekompozyty przetwarzają duże ilości materii organicznej, przekształcając ją z powrotem w nieorganiczną formę odżywczą.
Przykład:
Wspólny naziemny łańcuch pokarmowy detrytus to: Wypas łańcucha pokarmowego
Detrytus → Dżdżownica → Wróbel → Sokół


Food Web:
W naturalnych warunkach liniowe rozmieszczenie łańcuchów pokarmowych prawie nie występuje, a te pozostają połączone ze sobą za pośrednictwem różnych typów organizmów. Interlocking kilku połączonych łańcuchów pokarmowych nazywa się Food Web.
Sieć żywieniowa ilustruje kilka alternatywnych ścieżek. Sieci pokarmowe są bardzo przydatne w utrzymaniu stabilności ekosystemu. Jeśli liczba królików w danym obszarze maleje, oczekuje się, że sowy umrą z głodu.
Jednak ze względu na zmniejszenie liczby królików, więcej trawy jest pomijane, co pomaga zwiększyć populację szczurów. Teraz sowy żywią się szczurami i zwiększają liczbę królików. W ten sposób ekosystem nie zostaje trwale zakłócony, gdy żywność działa. Złożoność każdej sieci pokarmowej zależy od różnorodności organizmów w systemie.
W związku z tym zależy to od dwóch głównych punktów:
(i) Długość łańcucha pokarmowego:
Różnorodność organizmów w zależności od ich nawyków żywieniowych determinowałaby długość łańcucha pokarmowego. Bardziej zróżnicowane organizmy w nawykach żywieniowych, dłuższy byłby łańcuch pokarmowy.
(ii) Alternatywy w różnych punktach konsumentów w łańcuchu żywnościowym:
Bardziej alternatywami byłby wzór blokujący. W głębokich oceanach, morzach itp., Gdzie spotykamy różne typy organizmów, sieci pokarmowe są bardzo złożone.

Ekologiczne piramidy:
Ilościową i najłatwiejszą metodą badania relacji między organizmami w ekosystemie i pokazania ich w sposób schematyczny jest piramida ekologiczna, nadana przez Eltona (1927). W piramidach tych najniższy poziom troficzny jest tworzony przez producentów, podczas gdy najwyższy poziom troficzny jest taki, jak u zwierząt mięsożernych.
Zasadniczo rozważane są trzy typy piramid:

(i) Piramida liczb:
Piramida ta ilustruje związek między liczbą producentów, roślinożerców i mięsożerców. Organizmy danego obszaru są najpierw liczone, a następnie grupowane w ich poziomy troficzne. Przebadaliśmy trzy wspólne ekosystemy, a mianowicie. ekosystem leśny, ekosystem łąk i ekosystem stawu.
za. W ekosystemie leśnym kształt piramidy jest romboidalny. Producenci są reprezentowani przez duże drzewo o dużym kącie, od którego zależy kilka ptaków jedzących owoce itd. Dlatego liczba konsumentów pierwotnych jest większa niż liczba producentów. Następnie liczba odbiorców drugorzędnych i trzeciorzędnych stopniowo maleje.
b. W ekosystemie użytków zielonych trawą są producenci. Liczba konsumentów spada w kierunku górnej części piramidy. Liczba pierwotnych konsumentów lub zwierząt roślinożernych, takich jak szczury, króliki itp., Jest mniejsza niż liczba traw.
Liczba konsumentów wtórnych, takich jak jaszczurki, węże itp., Jest mniejsza niż liczba konsumentów pierwotnych. Liczba ostatnich lub trzeciorzędnych konsumentów jest nadal mniejsza niż liczba konsumentów wtórnych. Widzimy więc, że liczba organizmów stopniowo spada z pierwszego poziomu troficznego do ostatniego poziomu troficznego. Dlatego piramida liczby na użytkach zielonych jest prosta lub pionowa.
do. W ekosystemie stawowym liczba organizmów stopniowo spada z pierwszego poziomu troficznego do ostatniego poziomu troficznego. Dlatego piramida liczby w ekosystemie stawu jest prosta.

(ii) Piramida biomasy:
Całkowita masa organizmów nazywa się biomasą. Można go określić w kategoriach masy netto, suchej masy lub suchej masy bez popiołu. Biomasę w czasie pobierania próbek nazywa się stojącą biomasą lub stałą biomasą uprawną. W ekosystemie leśnym i ekosystemie łąkowym piramida biomasy jest wyprostowana. Ilość biomasy stopniowo zmniejsza się od pierwszego poziomu troficznego producentów do ostatniego poziomu troficznego zwierząt mięsożernych.
W ekosystemie stawowym liczba producentów jest duża, ale ich biomasa jest najmniejsza ze wszystkich, a ich rozmiary są bardzo małe. Ilość biomasy rośnie stopniowo wraz z pierwotnymi, wtórnymi i trzeciorzędnymi poziomami troficznymi. Dlatego piramida biomasy w ekosystemie stawu jest odwrócona.

(iii) Piramida energii:
Najbardziej idealną i podstawową metodą przedstawiania związków między organizmami w różnych poziomach troficznych jest piramida energii. Wiemy, że w każdym ekosystemie tylko producenci posiadają zdolność wykorzystywania energii słonecznej i przekształcania jej w żywność.
Energia w postaci żywności zostaje przeniesiona z jednego poziomu troficznego do drugiego. Dlatego przepływ energii jest zawsze jednokierunkowy. Ilość energii, która osiąga poziom troficzny netto jest mniejsza niż była obecna we wcześniejszym poziomie troficznym. W ten sposób ilość energii maleje wraz z każdym kolejnym wyższym poziomem troficznym. Dlatego we wszystkich typach ekosystemów taka piramida byłaby pionowa.

3. Cykl biogeochemiczny:
Transport i przemiany substancji w środowisku, przez życie, powietrze, morze, ląd i lód, są znane zbiorczo jako cykle biogeochemiczne. Te globalne cykle obejmują cyrkulację pewnych pierwiastków lub składników odżywczych, od których zależy życie i klimat ziemi.
Obieg węgla:
Ruch węgla w jego wielu formach, między biosferą, atmosferą, oceanami i geosferą.
za. Rośliny uzyskują dwutlenek węgla z powietrza i, poprzez fotosyntezę, inkorporują węgiel do swoich tkanek.
b. Producenci i konsumenci - przekształcają część węgla w żywności z powrotem w dwutlenek węgla poprzez oddychanie.
do. Dekompozyty - uwalniają węgiel związany z martwych roślin i zwierząt do atmosfery.
re. Inna ważna wymiana dwutlenku węgla zachodzi między oceanami a atmosferą. Rozpuszczony CO 2 w oceanach jest wykorzystywany przez florę morską w fotosyntezie.
mi. Dwa inne ważne procesy to spalanie paliw kopalnych i zmiana użytkowania gruntów. W przypadku spalania paliw kopalnych węgiel, ropa naftowa, gaz ziemny i benzyna są zużywane przez przemysł, elektrownie i samochody. Zmiana użytkowania gruntów jest szerokim pojęciem, które obejmuje szereg działań głównie ludzkich, w tym rolnictwo, wylesianie i ponowne zalesianie.

Globalny cykl węglowy jest niezrównoważony, co powoduje, że szybka globalna zmiana klimatu jest bardziej prawdopodobna. Poziomy CO2 w atmosferze rosną szybko, obecnie; są one o 25% wyższe niż przed rewolucją przemysłową. Dwutlenek węgla powstaje, gdy węgiel w biomasie utlenia się podczas spalania lub rozpadu.
Wiele procesów biologicznych uruchomionych przez ludzi uwalnia dwutlenek węgla. Należą do nich: spalanie paliw kopalnych (węgiel, ropa naftowa i gaz ziemny), rolnictwo typu "slash-burn", oczyszczanie ziemi na trwałe użytki zielone, grunty orne, osiedla ludzkie, przypadkowe i celowe spalanie lasów oraz niezrównoważone pozyskiwanie drewna i pozyskiwanie drewna z paliw.
Usunięcie pokrywy roślinnej z zalesionego hektara uwalnia dużo węgla z wegetacji do atmosfery, a także trochę węgla osadzonego w glebie. Pozyskanie drewna lub zrównoważone pozyskiwanie drewna opałowego może również obniżyć pokrycie roślinności i spowodować uwolnienie węgla netto.
Cykl azotu:
Prawie cały azot znajdowany w ekosystemach lądowych pochodzi z atmosfery. Niewielkie proporcje wchodzą do gleby w opadach deszczu lub w wyniku działania błyskawicy. Większość jednak jest utrwalona biochemicznie w glebie przez specjalistyczne mikroorganizmy, takie jak bakterie. Członkowie rodziny fasoli (rośliny strączkowe) i niektóre inne rośliny tworzą wzajemnie symbiotyczne związki z bakteriami wiążącymi azot.
W zamian za pewien azot, bakterie otrzymują z roślin węglowodany i specjalne struktury (guzki) w korzeniach, gdzie mogą istnieć w wilgotnym środowisku. Naukowiec szacuje, że fiksacja biologiczna na świecie dodaje rocznie około 140 milionów ton metrycznych azotu do ekosystemów.

Cykl fosforu:
Fosfor jest kluczem do energii w żywych organizmach, ponieważ jest to fosfor, który przenosi energię z ATP na inną cząsteczkę, napędzając reakcję enzymatyczną lub transport komórkowy. Fosfor jest również klejem, który utrzymuje DNA razem, wiążąc razem cukry dezoksyrybozy, tworząc szkielet cząsteczki I DNA. Fosfor wykonuje tę samą pracę w RNA.

Ponownie, kluczowymi źródłami pozyskiwania fosforu w systemach troficznych są i rośliny. Rośliny pochłaniają fosfor z wody i gleby do swoich tkanek, wiążąc je z cząsteczkami organicznymi. Po zażyciu rośliny fosforu są dostępne dla zwierząt, gdy konsumują rośliny.
Kiedy rośliny i zwierzęta giną, bakterie rozkładają swoje ciała, uwalniając część fosforu z powrotem do gleby. Raz w glebie fosfor może być przemieszczany od 100 do 1000 mil od momentu wypuszczenia ich przez jezdnie przez rzeki i strumienie. Cykl wody odgrywa więc kluczową rolę w przenoszeniu fosforu z ekosystemu do ekosystemu.
4. Sukcesja ekologiczna:
Stopniowe i ciągłe zastępowanie gatunków roślin i zwierząt innymi gatunkami, aż w końcu społeczność jako całość, zostaje zastąpione innym rodzajem społeczności. Jest to stopniowa zmiana i to właśnie organizmy, które wywołują tę zmianę.
Obejmuje procesy kolonizacji, zakładania i wymierania, które działają na gatunki biorące w nim udział. Występuje etapami, zwanymi stadiami seryjnymi, które można rozpoznać po zbiorach gatunków dominujących w tym miejscu w sukcesji.
Sukces zaczyna się, gdy obszar jest częściowo lub całkowicie pozbawiony roślinności z powodu zakłóceń. Niektóre typowe mechanizmy zakłóceń to pożary, burze wiatrowe, erupcje wulkanów, wycinki drzew, zmiana klimatu, poważne powodzie, choroby i epidemie szkodników. Zatrzymuje się, gdy skład gatunkowy nie zmienia się z czasem, a społeczność ta nazywa się społecznością klimaksową.
Rodzaje sukcesji:
Różne typy sukcesji zostały pogrupowane na różne sposoby na podstawie różnych aspektów.
Niektóre podstawowe rodzaje dziedziczenia są jednak następujące:
1. Podstawowa sukcesja:
Występuje na obszarze świeżo wyeksponowanej skały lub piasku lub lawy lub w dowolnym miejscu, które nie było wcześniej zajmowane przez społeczność żyjącą (biotyczną).
2. Sukcesja wtórna:
Odbywa się w miejscu, gdzie społeczność została usunięta, np. Na zaoranym polu lub jasno wyciętym lesie.
3. Sukcesja autogeniczna:
Po sukcesji sukcesji, w większości przypadków, to sama społeczność, która w wyniku reakcji ze środowiskiem modyfikuje własne środowisko, a tym samym powoduje, że nowe społeczności zastępują je. Ten przebieg sukcesji nazywany jest dziedziczeniem autogenicznym.
4. Sukcesja allogeniczna:
W niektórych przypadkach zastąpienie istniejącej społeczności spowodowane jest jednak w znacznym stopniu przez inne czynniki zewnętrzne, a nie przez istniejące organizmy. Taki przebieg określany jest jako sukcesja allogeniczna.
Na podstawie kolejnych zmian w zawartości składników odżywczych i energetycznych sukcesje są czasami klasyfikowane jako:
1. Autotroficzna sukcesja:
Charakteryzuje się wczesną i ciągłą dominacją organizmów autotroficznych, takich jak rośliny zielone. Rozpoczyna się w środowisku głównie nieorganicznym, a przepływ energii utrzymuje się w nieskończoność. Stopniowy wzrost zawartości materii organicznej jest wspierany przez przepływ energii.
2. Sukcesja heterotroficzna:
Charakteryzuje się wczesną dominacją heterotrofów, takich jak bakterie, promieniowce, grzyby i zwierzęta. Rozpoczyna się w przeważającym środowisku organicznym i następuje progresywny spadek zawartości energii.
Sukces ekologiczny oparty na siedliskach:
Znane są następujące rodzaje sukcesji, które są oparte na typie siedliska:
(i) Hydrosere lub hydrarch:
Ten rodzaj sukcesji występuje w zbiornikach wodnych, takich jak stawy, jeziora, strumienie itp.
Sukcesja występująca w zbiornikach wodnych nazywa się hydroserem. Jest to sukcesja występująca w środowisku wodnym. Rozpoczyna się od kolonizacji fitoplanktonu i ostatecznie kończy się w lesie. Istnieje około siedmiu etapów hydroseru.
1. Stadium fitoplanktonu:
To pionierskie etapy hydroseru. Na tym etapie występuje wiele organizmów, takich jak bakterie, algi i rośliny wodne. Wszystkie te organizmy dodają dużej ilości materii organicznej do śmierci i rozkładu.
2. Scena zanurzona:
Pojawia się po etapie fitoplanktonu, kiedy luźna warstwa błota powstaje na dnie stawu. Powstają pewne zakorzenione rośliny podwodne.
3. Pływający etap:
W miarę jak głębokość wody zmniejsza się, zanurzone rośliny ustępują miejsca nowej formie wodnej roślinności. Może to być przyczyną zniknięcia zanurzonych roślin. Później szybki proces glebotwórczy zmniejsza głębokość wody do tego stopnia, że staje się zbyt płytka, by przetrwać pływające rośliny.
4. Etap amfibii:
Ze względu na szybkie formowanie gleby stawy i jeziora stają się zbyt płytkie, więc siedlisko nie nadaje się do pływających roślin. W tych warunkach pojawiają się rośliny amfibii. Rośliny te żyją zarówno w środowisku wodnym, jak i powietrznym.
5. Etap Sedge-Meadow (marginalnie maty):
Powstaje gleba, co skutkuje podmokłą glebą, która może być zbyt sucha. Ważnymi roślinami tego stadium są członkowie cyperaceae i gramineae. Te suche siedliska mogą zupełnie nie nadawać się do hydrofitów, a zaczynają pojawiać się stopniowo krzewy i małe drzewa.
6. Scena leśna:
Na tym etapie w glebie gromadzi się duża ilość ludzi, bakterii, grzybów i innych. Wszystko to sprzyja wejściu wielu drzew w roślinność prowadzącej do etapu kulminacyjnego.
7. Etap Climax:
Hydrosere może zmienić się w klimatyczny las, roślinność. Na tym etapie najczęściej występują zioła i drzewa. Charakter kulminacji zależy od klimatu regionu. Jest to bardzo powolny proces i wymaga wielu lat, aby osiągnąć punkt kulminacyjny.

(ii) Xerosere lub xerarch:
Ten rodzaj sukcesji występuje w obszarach lądowych o niskiej wilgotności, np. Skałach, piasku itp.
Odbywa się na powierzchni, która jest wyjątkowo sucha, charakteryzująca się niedoborem wody i dostępnych substancji odżywczych. Rozpoczyna się na bazie skały. W tak ekstremalnie suchym środowisku mogą przetrwać tylko te rośliny, które są odporne tylko na ekstremalne suche środowisko.
Różne etapy Xerosere zostały opisane następująco:
1. Scena z liszajowca:
Skały są całkowicie pozbawione wilgoci i składników odżywczych. To pionierzy w Xerosere. Ważnymi porostami skorupiaków są Rhizocarpus. Porosty wydzielają kwas węglowy, który pomaga korodować i rozkładać skały, uzupełniając inne czynniki związane z mieszaniem się.
2. Foliose Lichen Stage:
Wietrzenie skał i rozkład gnijących porostów tworzy pierwszą warstwę gleby na powierzchni skał. Stopniowo warunki stają się korzystne dla istniejących porostów foliozy i fruktozy.
3. Moss Stage:
Po etapie porostów foliozy i fruktozy następuje stadium mchów. Wraz z formowaniem gleby na powierzchni skał, masy kerofityczne rosną i stają się dominujące. Typowymi przykładami mchów ksenofitycznych są: polytrichum, tortula, grimmia itp. Ta mata z mchu powstaje na ziemi. Mata staje się grubsza, co zwiększa zdolność zatrzymywania wody w glebie. Teraz stadium mchu zastąpiono nową rośliną zielną.
4. Scena zielna:
Początkowo niektóre roczne zioła migrują i kiełkują. Ludzie z gleby zwiększają się z roku na rok, ponieważ z powodu śmierci i rozkładu rocznego zioła. Powoli rosną biennale i wieloletnie zioła. Im więcej materii organicznej i składników odżywczych gromadzi się w glebie. To sprawia, że siedlisko jest bardziej odpowiednie dla roślin drzewiastych.
5. Stopień krzewów:
Coraz więcej gleby powstaje w stadium trawiastym dla drzewiastych krzewów. Zioła są zacienione przez rosnące krzewy, gnijące zioła i liście, gałązki krzewów. Wzbogacają one również glebę w próchnicę. Wilgotność wzrasta w takich obszarach. Wszystko to sprzyja rozwojowi dużych drzew mezofitycznych.
6. Etap Climax:
Ten etap jest zajęty przez dużą liczbę drzew. Pierwsze drzewa rosnące na takich obszarach są stosunkowo niewielkie wraz ze wzrostem zdolności zatrzymywania wody w glebie, drzewa te znikają i rozwijają się duże drzewa mezofityczne.

(iii) Lithosere: Ten rodzaj sukcesji rozpoczyna się na gołej skale.
(iv) Halosere: Tego typu sukcesja rozpoczyna się na wodzie słonej lub glebie.
(v) Psammosere: Ten rodzaj sukcesji rozpoczyna się na piaszczystym obszarze.
Proces sukcesji ekologicznej:
Każda podstawowa dziedziczność, niezależnie od odsłoniętego obszaru, z którego inicjuje, wykazuje następujące pięć kroków następujących po sobie:
(i) Nudacja:
Etap obejmuje rozwój gołego obszaru, który może być spowodowany erozją gleby, osadzaniem itp.
(ii) Inwazja:
Krok polega na udanym założeniu gatunku na gołym terenie. Gatunek dociera do tego obszaru z innego regionu.
(iii) Konkurencja i wspólna akcja:
Gatunek zajmujący nowy obszar rozwija wewnętrzną i między-specyficzną konkurencję o żywność i przestrzeń kosmiczną. Zakończenie między istniejącymi gatunkami i tymi, które właśnie wkroczyły na obszar, powoduje zniszczenie jednego z nich, który jest nieodpowiedni.
(iv) Odczyn:
Gatunek lub społeczność, która utworzyła się w nowym obszarze, wpływa na środowisko, modyfikując światło, wodę, glebę itp. Powoduje to eliminację społeczności, która następnie ustępuje miejsca innej społeczności, dla której bardziej odpowiednie jest zmodyfikowane środowisko. Różne społeczności lub etapy reprezentowane przez połączenie mchów, ziół, krzewów i drzew zastępujących się nawzajem podczas sukcesji są znane jako stadia surowicy, społeczności surowcowe lub etapy rozwojowe.
(v) Stabilizacja:
Jest to ostatni etap, w trakcie sukcesji, kiedy społeczność osiąga równowagę z klimatem danego obszaru i staje się względnie stabilna. Ta ostatnia społeczność znana jest jako społeczność klimaksowa.